这是镜像神经元相关作品的读书笔记,主要内容是《镜像练习》,这本书介绍现代神经科学前沿的镜像神经系统研究,并告诉我们这对我们的生活、对我们子女的教育、对我们的工作、对伴侣关系中的相互交往,以及对社会和政治生活意味着什么。最重要的是 —— 我们能更好地了解自己,并认识到良好的自我照顾意味着什么。
历史
镜像神经元的研究基于前人建立的科学基础,了解发现的过程有利于理解这一系统。
古典阶段
笛卡尔的身心二元论将身体和灵魂分离,为生理学奠定了基础。对身体的研究可以独立于精神,生命机制从神秘主义中分离出来。
谢灵顿在纯粹的身体研究中,对脊髓反射的研究奠定了神经生理学基础,某种刺激有极高的可能性造成某种特定反应。巴甫洛夫的条件反射让人类意识到,一种反应紧随一种刺激,而且是可预测的。视觉反应不能机械地刺激唾液分泌,其中有某种精神过程,这代表心理过程也是可研究的。 笛卡尔分离的身心在启蒙运动中又合二为一。
启蒙运动中,洛克和休谟认同感觉论:一切源自经验,不存在缺乏经验基础却能出现于知识中的思想。外部刺激在起作用时会与我们的身体进行接触,然后重新组织和呈现。精神是一张白板,精神仅仅是各种感觉的叠加或集合,有了经验,知识才能产生,经验则通过我们的感觉被察觉、整理,人类是经验的集合。
神经科学家在这种思想指引下,由下至上,从每一个细微结构逐层研究神经系统,认为是从每一个系统的混合、关联中诞生了感知和认知。
古典阶段由下至上的研究明晰了人体结构,为语言学阶段奠定了基础。
语言学阶段
乔姆斯基从语言领域出发,提出人类天生存在语言系统,学习走路和学习骑自行车是两种概念,一些系统成熟之后,其功能自然生效,外部刺激虽然重要,但只是起到促使我们的天然机制运转起来的作用,并不是经验主义认为的,我们是依靠感受的复杂组合来理解世界和他人的。我们不是白板,并不会从外界感受到什么就在上面写什么,我们的白板上早已提前写下了很多东西,但只有光线(即经验)照在上面时才能显现。
斯金纳则提出,我们的行为都建立在刺激-反应关系基础上,如果一项行为的效果是积极的,就会被重复,如果是消极的,就会被消除。起决定性作用的是效果,行为单纯建立在这种强化机制上,语言不具有特殊性。
乔姆斯基则坚持语言天生,人类具备计算机制的语言功能器官,能够以递归的形式运行,逐渐制造出非常复杂的结果,机制之间相互交织、相互嵌套。斯金纳和乔姆斯基的争论核心是经验在神经系统的研究中起到什么作用。
现代阶段
1959年,休伯尔和威泽尔发现,直接用光点照射视网膜,视觉皮层神经不会兴奋,视觉皮层抑制区会阻止兴奋区的兴奋。视觉神经元的兴奋,需要一个有完整结构的视觉刺激。他们在后续研究中发现,一些神经元会对竖直移动做出反应,一些会对水平移动做反应,一些会对椭圆移动作反应,他们的研究发现了神经元分解物体来进行视觉描绘的机制,休伯尔和威泽尔因此获得1981年诺贝尔生理学或医学奖。
科学家格罗斯和罗卡·米兰达偶然观察到,当他们把视线转向一只猴子时,猴子的颞叶神经元竟然兴奋了,他们马上意识到,引起猴子神经元兴奋的刺激其实是人的脸孔。感光神经-视觉皮层-颞页的机制在相关研究中被发现了,刺激被感光神经接收,在视觉皮层被分析后,信息在颞页综合起来,颞页描绘事物各部分的不同环节通过颞页神经元的连接相互补充,然后解读出事物,通过fMRI(功能性磁共振成像),他们证实了面孔识别区在人脑中的存在。
科学家井上达二发现了视觉系统的生理结构,左侧视觉刺激会引发右脑兴奋,产生对左手的控制活动,视觉系统和运动系统形成左右交叉的配合。
这些研究基于经验主义,却发现了天生器官。
镜像神经系统的发现
1988年,贾科莫·里佐拉蒂等神经科学家在意大利帕尔马大学开展了一项实验,研究恒河猴前运动皮层神经元控制抓握动作的基础性研究。
他们首先发现,运动区很多神经不会在简单运动中兴奋,只有在为了达到某一目的才进行的运动中才会被激活,一些区域是“抓取”,一些区域是“持握”,这构成了"运动词典“,大脑容器里已经写好的我们全部运动的指令集。
我们并不是“学会“走路,是儿童在发育过程中一步步实施天生系统写好的行为,通过与现实接触逐渐熟练和精细化,人们不是用学会弹钢琴、下棋的方式来 “学会” 走路的,同一个词语描绘了两种不同的行为。
💡 我的理解,像呼吸、走路、说话都是人类大脑指令集已经预设,具备肺、腿、喉等肉体硬件的,我们之所以能 “学会” 走路、说话,是因为我们本来就会,这里的 “学会” 是硬件 “激活” 的过程,或者说随着儿童发育,脑指令集(相关功能区)和躯体硬件调整适配的过程。骑自行车、写字、弹钢琴等 “学会” 则需要自行车、笔、钢琴等外置硬件,这类 “学会” 是基于原始指令集复杂叠加的模仿,是基于硬件的软件层行为,不断练习刺激的神经元突触连接就是软件的写入。
在恒河猴前运动皮层神经元控制抓握动作研究中还发现了近身空间,只有当我们有能力移动时,才能获得关于空间的概念,远和近在人脑中不是一个空间整体,而是被定量划分成两个空间。规范性神经元—— 负责解读物体大小、形状的视觉-运动神经元,能够辨识物体并将其内化,将物体形状大小转化为与之相适应的动作来进行抓取。
这些帕尔马大学的科学家由于经费问题和行政审批问题进度缓慢,贾科莫·里佐拉蒂在一次国际会议上和在做相似研究的日本科学家酒田英夫相遇,他们都在研究物体可供性(Affordance)——物体物理性质的集合,能让个体理解其可用性。当我们抓起咖啡杯时,规范性神经元解读物体的可供性,根据物体的形状和大小转化为行为,从指令集中挑选出有效的抓取方式。
酒田英夫介绍他们加入日本的人类前沿科学计划(HFSP),这个计划给他们提供了大额资金,深入研究猕猴的运动皮层会对视觉刺激做出反应的工作原理。他们有两个选择:1. 正统研究方法,引导猴子进行运动,在此基础上观察 2. 自由观察,让猴子自由活动,观察什么时候神经元会兴奋。
选择非正统研究方法,很快带来了两个出乎意料的发现:
- 一些神经元竟然在猴子观察研究员取食时兴奋了
- 当猴子做相同的取食活动时,同一区域的神经元兴奋了
他们在实验中证实,一些神经元既会在猴子完成动作时激活,也会在猴子观察学者做相同动作时激活,这些神经元被命名为 镜像神经元(mirror neuron)。
镜像神经元概述
观察即理解
镜像神经元是动物在执行某个行为以及观察其他个体执行同一行为时都发放冲动的神经元。
无意识中无时无刻不在运行的镜像神经元系统有一个重要特性 —— 观察即理解。用心理学名词来说就是自他同一视,即 “感同身受”。这种感觉-运动神经系统在动作观察和动作执行两个阶段皆被激活,可能是模仿的基础系统。
镜像神经元改写了传统观念,过去我们认为模仿的过程是:观察→认知→动作,现在的发现是:观察[运动皮层-模拟运行]→认知→动作,正是因为有运动系统的模拟运行,我们才能理解他人的行为 —— 就好像我们自己也进行了该行为一样。
模仿
动作和模仿是人类发展的基础,其次用语言来丰富并巩固各种感觉。通过镜像神经元,我们能通过想象自己亲自体会的方式来 “阅读” 他人的行为,用这种方式达成人与人之间姿态与行为的共享。我们在观察他人时会将他人置换成自己,就好像在镜中注视自己。
简单的模仿只需要跟着看到的去做就行了,将一种视觉表现转化为动作表现,这种动作表现精确的再现了从视觉表现中解读出来的数据。模仿基于感知与行为之间共通的表现域来实现。感知与行为之间拥有同一种机制:观察即理解。
模仿学习有两个阶段:第一个阶段中,观察者将需要模仿的动作切分为基础运动行为,这些行为在此前已经存在于自己的储存目录中(神经元连接)第二个阶段,观察者会根据不同的任务来重新组织这些运动行为,产生新的排列顺序或是非顺序的新模式。
这两个阶段的生物学基础是同一套感觉-行动神经系统,在第一阶段中,观察者在观察时,模拟运行被观察者做出的动作,但不输出为行为;在第二阶段中,观察者产生第一阶段相同区域的神经兴奋,输出为行为。
模仿实验
《镜像练习》中的一个例子:一些之前从未学过吉他的学生需要观察一位吉他教授弹奏几个和弦,然后一组要重复和弦,一组要完成另一项完全不同的运动,用电位监测工具监测脑部活动。实验分为四个阶段
- 观察教师演奏和弦
- 学生为弹奏和弦做准备
- 和弦演奏
- 休息
在观察时,两组学生的顶叶-颞叶回路都出现激活现象,通过镜像机制,看到的动作转化为可能的行为。但在准备阶段中,要进行和弦演奏的学生镜像回路保持了激活状态,上额叶 46区(工作记忆)保持兴奋;要完成另一项完全不同的活动的学生,在观察阶段中激活的区域兴奋消失。
语言的习得
运用到语言学上,我们之所以能理解音素,是因为我们也能重复相同的发音,当一个男人/女人/小孩/计算机发出一个”biu“时,声调的差别很大,我们为什么认为是相同的音节呢? 因为这些 “biu” 激活的,都是我们自己用来发出这个音节时使用的运动系统。决定一个声音语音学特性的是其特殊的发音方式,即器官的运动。
人们模仿眼睛看到的发音方式,并与耳朵听到的发音建立联系。模仿与语言在度过了最初的磨合阶段后开始并肩前进,彼此交叉、彼此巩固。
幼态持续和人化过程
人类和猴子的胚胎起始阶段相似,但在分化阶段人类胚胎会保持更长时间,人类发育是滞后的,宫内周期长,出牙晚,极晚才脱离父母独自生活,这是一种幼态持续。
婴儿出生后就会将手伸向一个物体,相信自己能抓住它,然后遭遇失败,运动行为本身就带有目的性,只是需要与现实进行接触逐渐精细化。这种 “学习” 需要时间,一个器官的发育,一种功能的完善都需要教育和练习。
婴儿发育迟缓,但大脑已经预置了“抓取”命令,神经元确实执行了,但神经通路还不能很好的运转,动作就显得不精准,而且,如果一位成年人试图完成某种行为却连续遭遇几次失败,他就会停止(斯金纳模式),但儿童不会,儿童能够持续几个小时不停的尝试抓取物体,就好像有一种“本能”驱使他们要学会这件事。
就像经常演奏钢琴的人大脑皮层对应手指活动的区域更为发达,学习抓取物体的婴儿身上发生着类似的事情。
幼态持续缘于人类是“早产”的,马出生一个小时就会跑,小鸡破壳不久就能自行觅食。人类婴儿只会哭,12-14个月才会摇摇晃晃的站立,18-21个月才达到黑猩猩幼崽出生时的成熟度。
小鹿从被生下就是鹿,小鸡从鸡蛋里破壳而出就是鸡,婴儿则并不是同一概念下的 “人”,一个婴儿更像是在母体外发育的胚胎,在母体外通过练习完成其他动物在子宫里的发育过程,婴儿的多个神经系统经历数年才发育完整,这个母体外的练习过程可以称为人化过程。
这种出生时的不完善带来了人类的巨大可塑性。
模仿的社会意义
人类之所以成为人类,正是因为我们频繁模仿自己的同类。
通过模仿进行学习的能力是文明得以发展的关键因素,模仿构成了外部记忆,一个个体制造工具的能力可以在群体中传播,如果没有模仿,创造力也就没了意义。复制同类的行为标志了两个个体之间的相似性,这种复制行为在两个人之间指明对应关系 —— 我和你一样,将自己的经验和他人的经验进行共享、将自己融入他人。
我们常以为其他灵长类动物有和人类相同的模仿能力,但灵长类动物的模仿只是一种效仿(emulation)。效仿基于目的,不精确模仿过程。人类儿童在一岁时就有能力模仿成人,他们会精确地重复看到的动作。灵长类动物再现它们观察到的动作的目标,并不完成动作本身。
人类的镜像机制更复杂,也能够完整重复不及物动作,或缺少明确目标的直接及物动作。
💡 实验中的恒河猴,看到研究人员抓握香蕉时产生兴奋,看到研究人员抓握空气时不兴奋。这两个动作的视觉刺激引发的视觉神经兴奋是相同的,但猴子并不产生运动神经兴奋(模拟运行)来理解非及物动作,而人类的运动神经对看起来无意义的动作也进行模拟运行。这可能是由早产带来的神经系统不完善(相对于猴子)形成的,对于猴子,幼态持续和时刻运行的毫无目的的运动神经兴奋是一种基因缺陷,徒费营养和能量。但对于人类,理解无目的动作的缺陷让我们创造出更为复杂的行为,大脑也越来越大,最终成为了今天的人类这个物种。
💡 观察即理解(模拟运行),但人类也会像猴子一样,把对象视作一串简单动作进行忽视。当我们的注意力在手机或正思考的问题上时,路边行人、舞者被我们一般化为在行走、在动。但当我们的注意力集中在行人身上,漂亮的礼服会引发我们模拟运行自己穿上类似衣服的情境,啊她一定是去约会吧。舞者毫无意义的(对于猴子)用头在地上转圈圈的行为,引起我们模拟运行正在做这么酷的动作,有种想要跟着他一起舞动的运动冲动。
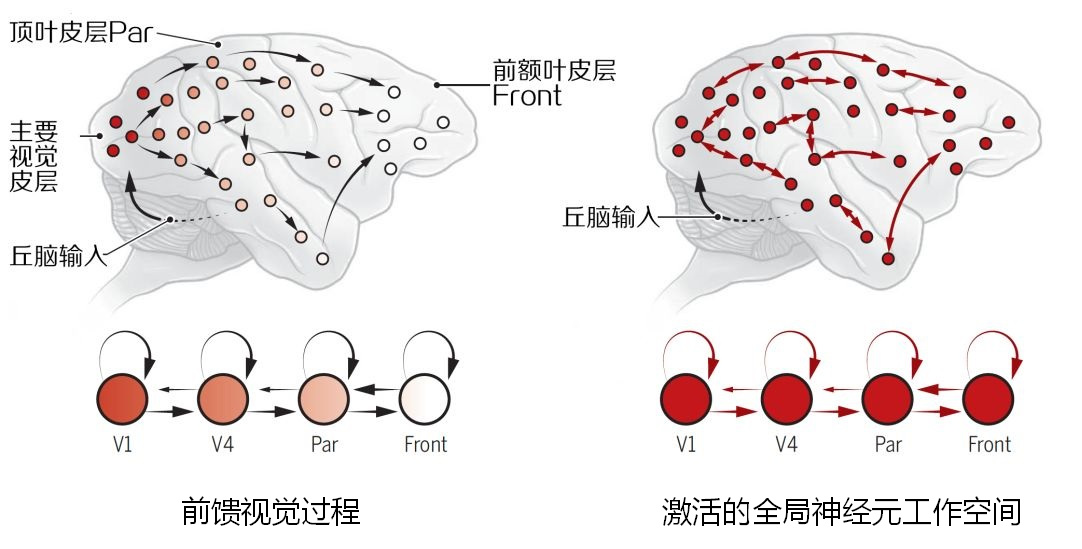
💡 这里同样有阈值的概念,视觉信号引起的过程兴奋是不可用于的(不可调节),见左侧图示。当刺激强度、持续时间满足条件时,会激活强的、持续的前额叶皮层活动,网络回响既允许视觉在意识上得到表征,又可用于其他认知系统,见右侧图示。强的、持续的额叶皮层和其他系统的相互作用使信号成为一种自我维持的表征,大脑在观察动作的同时,运动系统在进行运动模拟,只是不输出为动作,这种模拟的大脑神经元的兴奋也是记忆的生物基础,兴奋过后的神经元连接如同计算机中被改写过的 01存储,让我们记得一连串的动作。(镜像并不仅仅是视觉刺激的结果,其他感觉也可以引发模拟运行)
快乐原则(Pleasure Principle)
斯金纳认为模仿他人是因为我们有欲望得到他人所渴望的东西,但并不总是这样。成功的动作模仿当然是鼓舞人心的,如果我看到有人做了什么,然后我也成功的做到了同样的动作,我就会体会到一种快乐。但为我带来快乐效果的是我自己的动作,而不是另一人决定的那个动作。
意识
意识起源 过短/被紧随其后的另一个刺激遮掩的刺激只停留在视觉区,不会激活额叶。动物更多的是选择性注意,聚焦一个刺激,有效处理少量刺激。人类的优势是,能够让整个大脑都参与刺激的处理和反应过程,用分解的方式对刺激做出反应,处理刺激的方式让我们的大脑变大。双手和语言促进了这一过程。这种连续、有序的神经兴奋可能是意识的起源。
自由意志存在吗?一只老鼠绝不会只会寻找一处观赏夕阳的好位置而到处游走,所有生物都基于生理需求进行活动:吃、喝、繁衍、好奇心。人类有着这之外的浪漫行径,使得人类看似存在自由意志,但这可能只是对无目的行为镜像,理解无目的动作的大脑 “缺陷” 的一个副产物。
人类有自由意志的可能性很小,从物理上讲,人类不是什么特例,但从哲学的角度,当你在内心决定如何行动的时候你是自由的,当别人代替你的位置选择你需要做什么的时候,你就是不自由的。
规范性神经元
规范性神经元和近身空间是相关研究的成果。
规范性神经元能够辨识物体并将其内化,负责解读物体可供性(Affordance,物体物理性质的集合,能让个体理解其可用性),物体是按照其 “可处理” 的方式呈现的,抓住三维物体的方法有一种或多种,这些方法并不是无穷的 —— 如果我想喝一杯咖啡,只有一种方式让我抓起咖啡杯,否则我就可能让咖啡杯掉落或将咖啡洒出去。规范性神经元解读物体的可供性,然后将其转化为一种运动行为 —— 回到了运动词典 —— 词条说 “抓取”,规范性神经元就从词典中挑选出适合的行动,根据物体的形状和大小进行调整。
部分抓取动作是先天的,猕猴具备三种抓握物体的基本方式:全手抓握(全部手掌用力抓握)、精细抓握(用食指和大拇指抓取)、手指抓握(用全部手指进行抓取)。其他抓握方式也存在,但最基本的只有这三种,其他抓取方式中一些是后天学习的结果。
近身空间:空间感不像我们直觉上认为的那样是一个整体,而是被定量的划分成两个空间。近身空间会呈现在运动区域里,其形成源于运动。(庞加莱:只有我们有能力移动时,才能获得关于空间的概念,空间不是康德所认为的先验概念)
空间想象忽略症:患者会忽视视觉区一侧物品的物体可供性,症状比如吃一盘意面时只吃右边,尽管他看到了左边的意面,他能准确的描述左边的意面,他知道左边的意面可以吃,但大脑不产生吃左边意面的兴奋,就好像左边的意面是一个不及物的存在。
疾病感缺失症:患者会感到持续或反复的消失自我感或者失去现实感或者二者兼而有之。患者感到自己从心理过程、身体中分离出来,有一种“自动化感”,经历着生活改变,但却没有任何体验或参与感,仿佛生活是在电影里的场景。
镜像神经系统在哪里
镜像神经系统位于前运动皮层、后顶叶、颞上沟、脑岛等位置,这一系统不是一个新发现的和其他功能区明确区分的区域,而是感觉、运动神经等系统关联起来的一个包括语言、动作、识别、模仿的功能性概念。
对一个行为的理解是视觉、智力、模仿(镜像)的共同结果。没有运动系统参与的简单视觉感知只是对运动的视觉层面描述,不能为该行为的内在组成提供精确的信息,而这些信息是我们理解行为本质,即行为的目的与实现方法的重要一环。正是因为有了运动系统在观察时的模拟运行,我们才能理解他人的行为,就好像我们自己也进行了该行为一样。
体素
体素(voxel)被作者定义为会引发核磁共振图上的相同位置产生兴奋的刺激。体素是共情、暗示的基础。相同的体素会在 A 表达情感时,引发观察者 B 在相同区域兴奋。例如 A 闻到臭鸡蛋味道表达恶心,B "读到" A 的表达,和 A 觉得恶心时兴奋的相同区域产生兴奋。生物机制决定我们一定会成为别人的一部分,别人也一定会成为我们的一部分,这种同感是社会、文化的基石。
自闭症
可以用一个简单的动作过程理解自闭症:拿起巧克力。
普通儿童在拿起巧克力的动作时,张嘴用到的肌肉就有电位反应,自闭症儿童直到放入口中的动作时,张嘴用到的肌肉才会活动 —— 普通儿童能够打出很大的提前量来计划行为,自闭症儿童知道自己要什么,但意图不会转化为连续的运动行为,而只是在完成一个个单独的动作(行动)。
碎镜
自闭症儿童对实验人员拿起巧克力放入口中的行为,也未激活张开嘴巴需要的肌肉。对于普通儿童完整的镜像对象,对于自闭症儿童如同一面破碎的镜子,完整的动作是破碎、无组织的。他们理解不了他人的行为以及自我的内部意图,只能对内部之外的思考进行理解。
总要解读别人行为的人生是艰难的,他们的每一次思考都是一道谜题。在这些困难面前,受自闭症影响的儿童选择抽身事外,与周围世界或环境分离。镜像神经系统功能受损/发育不良又导致了自闭症儿童的严重认知障碍。
空堡垒
如果存在一个你不能理解的世界,如果你不能理解他人的意图,你就会发展出不关心他人的态度,将自己关在某些确切的事物中能让你觉得更有保障,从而摆脱困难、尴尬甚至可怕的情况。


